Photon Counting Detectors
High-performance SPAD, PMT and hybrid detectors for reliable photon counting across materials science, life science, quantum optics, and metrology.


Simplify your materials characterization with one flexible TRPL microscope enabling multiple methods for precise and efficient analysis.

Complete confocal fluorescence microscope that empowers researchers to advance quantitative functional imaging from individual molecules to cells and tissues.

Compact FLIM and FCS upgrade kit that adds advanced functional imaging and correlation analysis to existing laser scanning microscopes.

Designed for flexible, sensitive, and precise steady-state and time-resolved spectroscopy across the UV to NIR range and time scales from picoseconds to milliseconds.

Modular lifetime spectrometer designed for flexible fluorescence and photoluminescence measurements in both materials and life science research.

Add spectral and time-resolved photoluminescence to your setup through flexible microscope–spectrometer coupling options.

Get the most out of superconducting nanowire detectors in large-scale quantum communication and computing experiments requiring precise multichannel timing.

Boost your time-resolved experiments with a flexible, high-precision time tagging and TCSPC unit for materials science and quantum sensing.

Scale your photonic quantum computing and detector characterization setups while maintaining performance, flexibility, and high data throughput.

Compact 3-color picosecond laser delivering flexible ns to ms excitation with cost-effective multicolor performance and straightforward operation.

Smart picosecond laser diode heads covering UV-A to NIR, providing the right combination of power, pulse width, and diode type for any time-resolved technique.

VisUV provides clean short pulses and stable timing across key UV and visible wavelengths, including deep UV lines as well as 488 nm and 532 nm.

Enhance your single-photon counting experiments with wide dynamic range and excellent timing precision in the UV and visible even at the highest count rates.

Capture even the weakest signals over large areas with maximum dynamic range and enhanced low-light sensitivity in a compact detector design.

Unlock spatially resolved single-photon detection with a 23-pixel SPAD array, combining low dark counts and precise time tagging for advanced experiments.
Advanced FLIM analysis software for fast, accurate interpretation of lifetime imaging data.
Intuitive, free software solution for real-time, high-precision photon data acquisition, visualization, and initial data analysis.
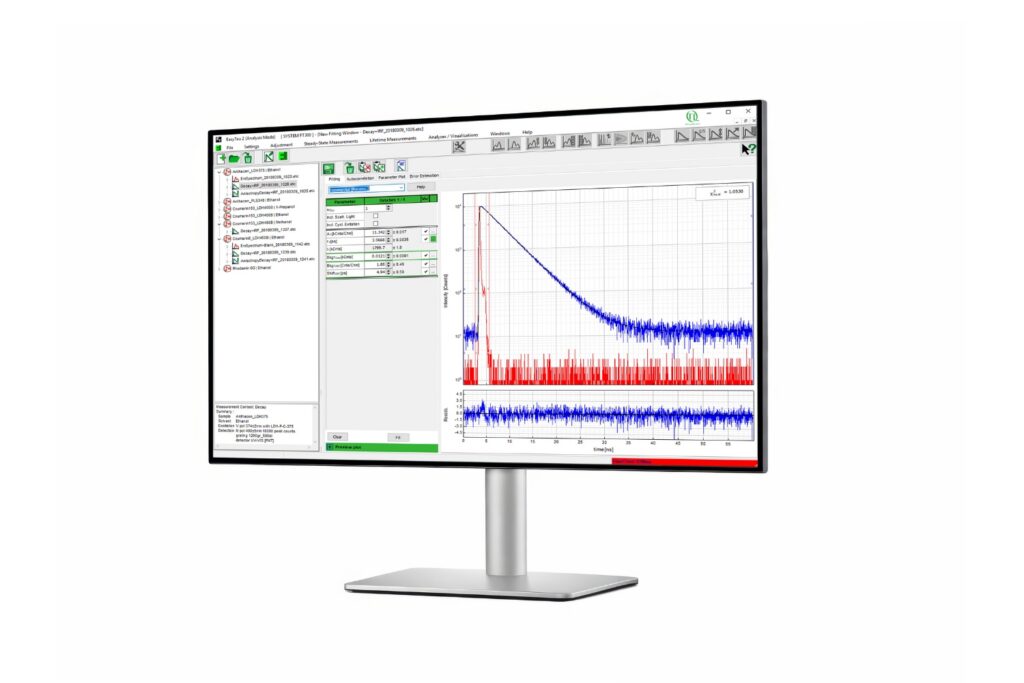
Advanced software for time-resolved fluorescence acquisition and analysis.
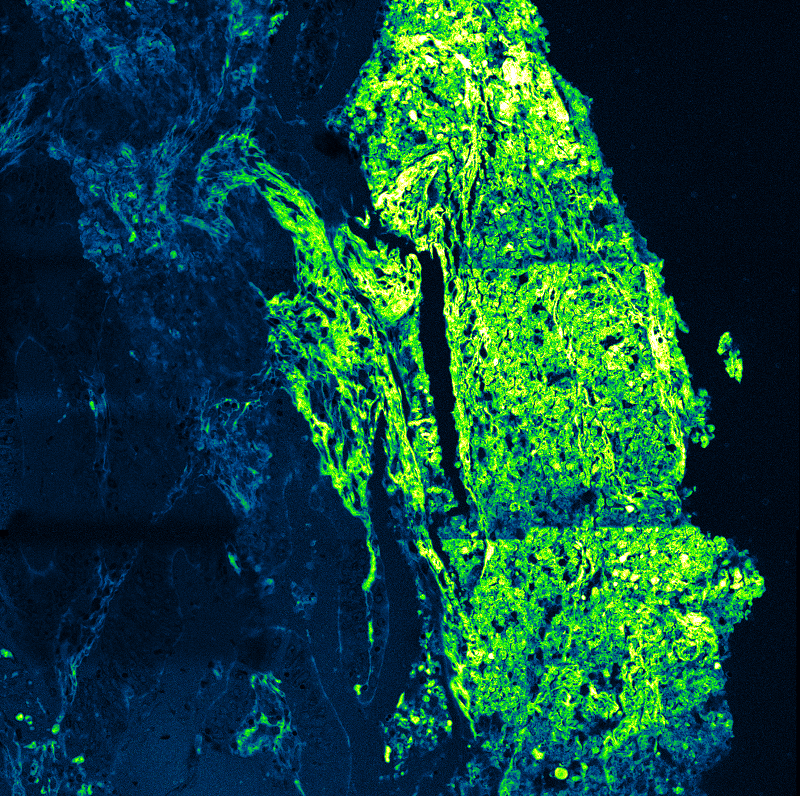
An imaging technique that uses fluorescence lifetimes to generate image contrast.
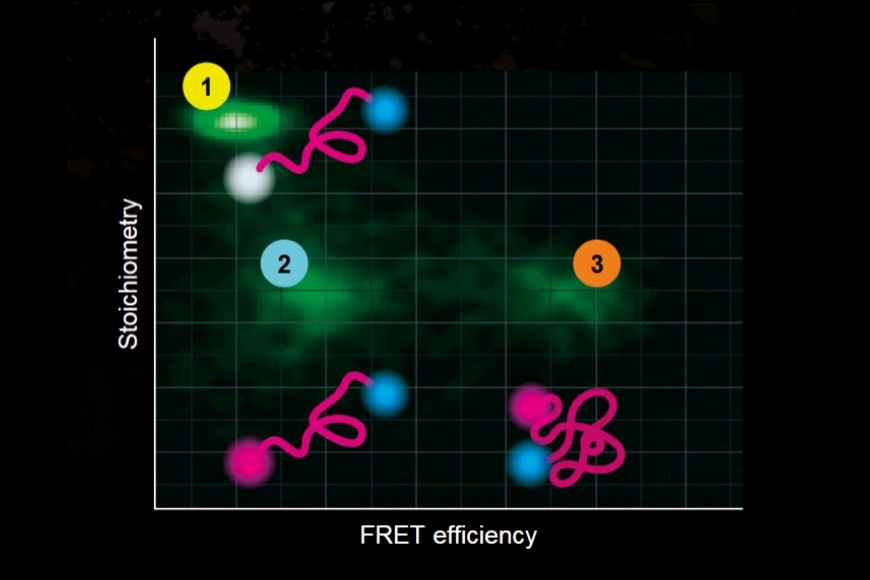
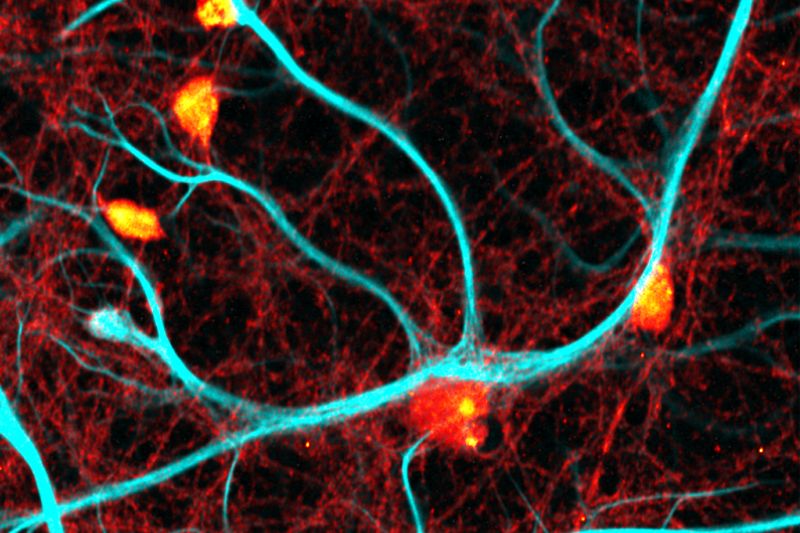
Investigating how proteins dynamically explore multiple conformational states that control biological function.
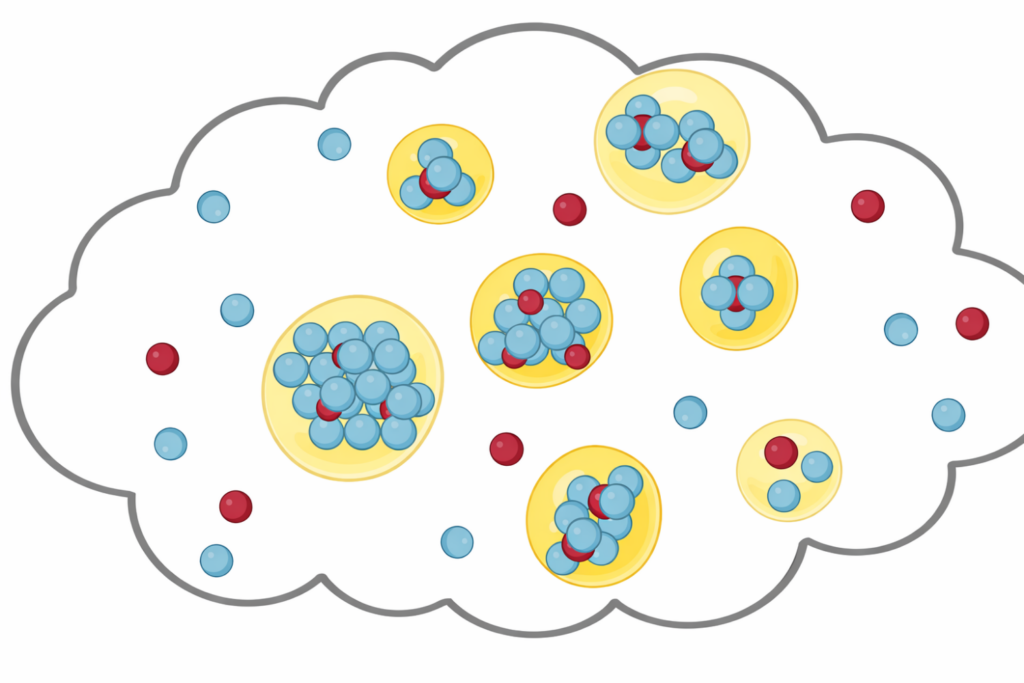
Investigating how biomolecules separate into dynamic liquid phases to organize cellular space and regulate biological function.
A time-resolved technique that measures photoluminescence lifetimes to reveal excited-state dynamics in materials.
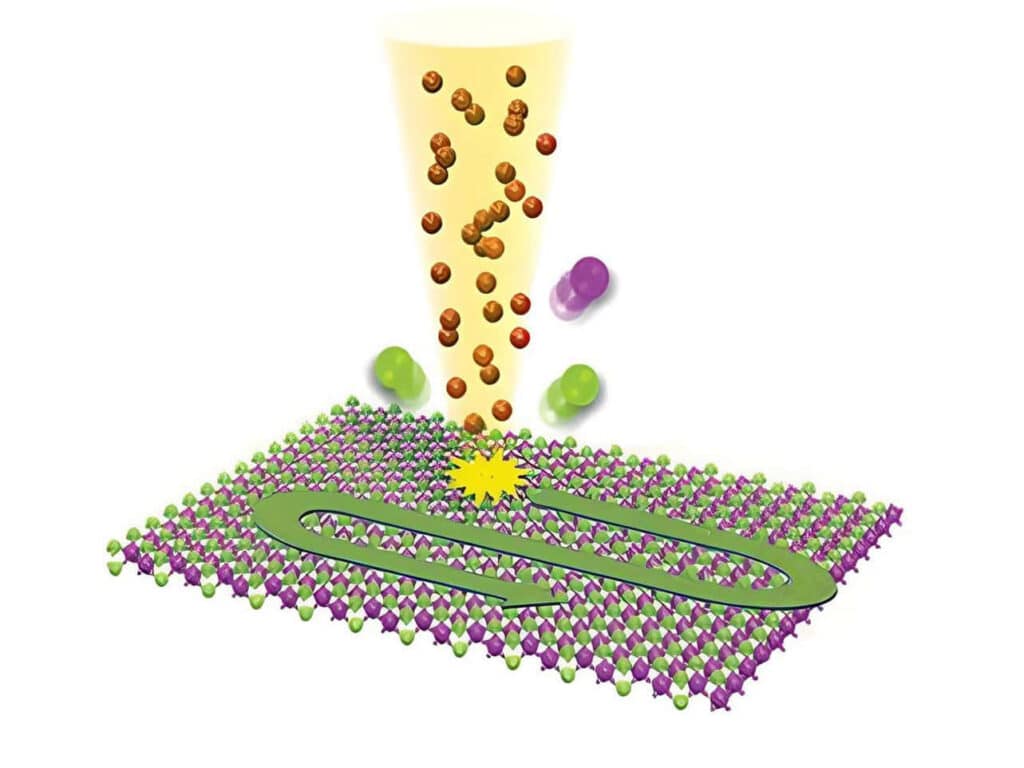
Studying exciton dynamics, charge carrier processes, and structural properties through optical and time-resolved characterization methods.

Investigating charge-carrier lifetimes and recombination dynamics to enable precise optical characterization of material quality and device performance.
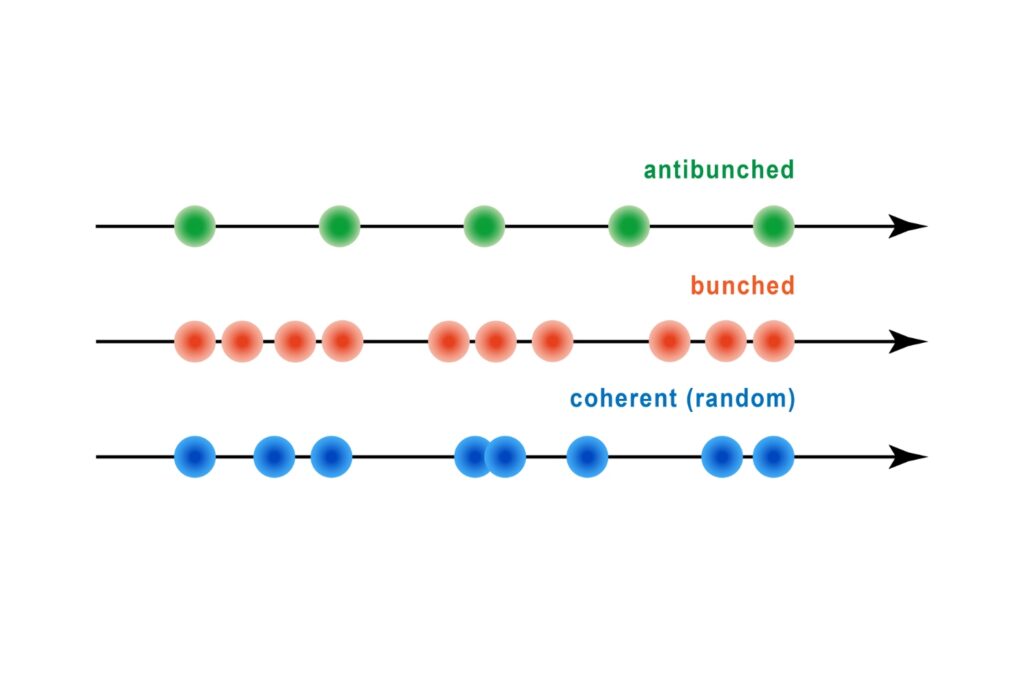
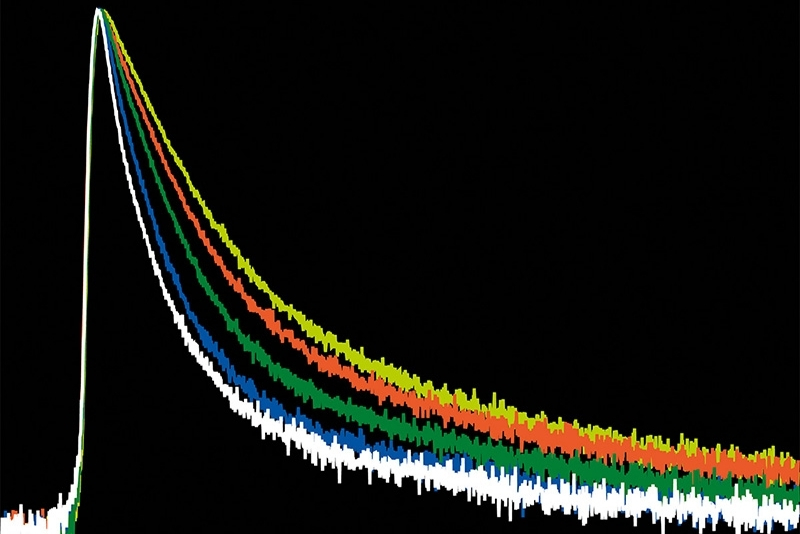
A quantum optical signature revealed by time-resolved photon correlation analysis to identify single-photon emission in materials and nanostructures.
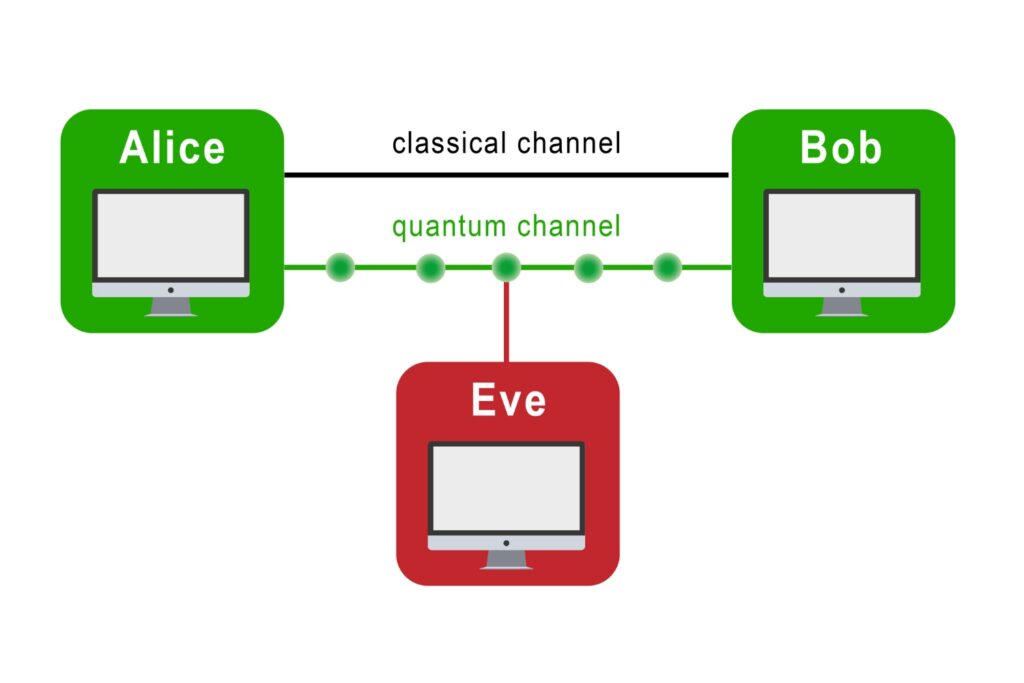
The transmission of information using individual photons, using quantum effects to ensure absolute security.
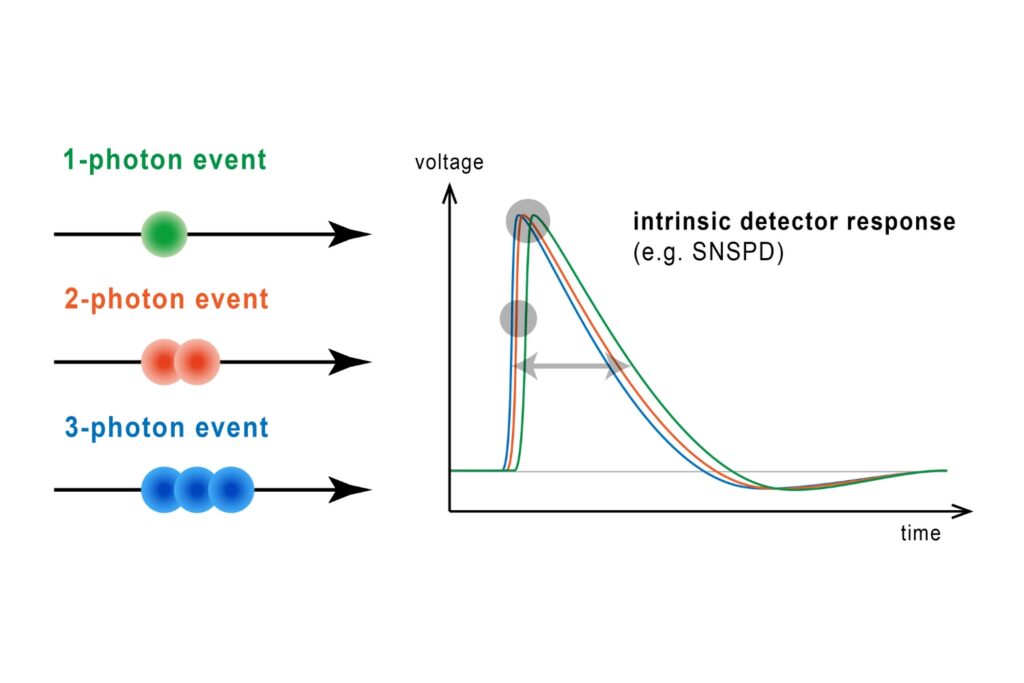
Quantifying photons per detection event enables direct access to photon-number statistics, providing insight into quantum and statistical properties of light.

An optical technique that analyzes light emission under electrical excitation to reveal electronic properties of electroluminescent materials.
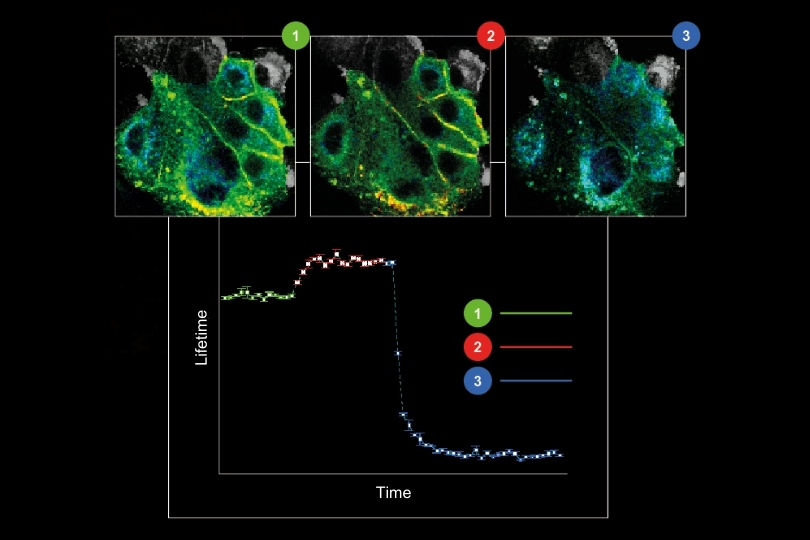
Monitoring environmental signals and trace compounds to understand dynamic changes in natural and engineered environments.
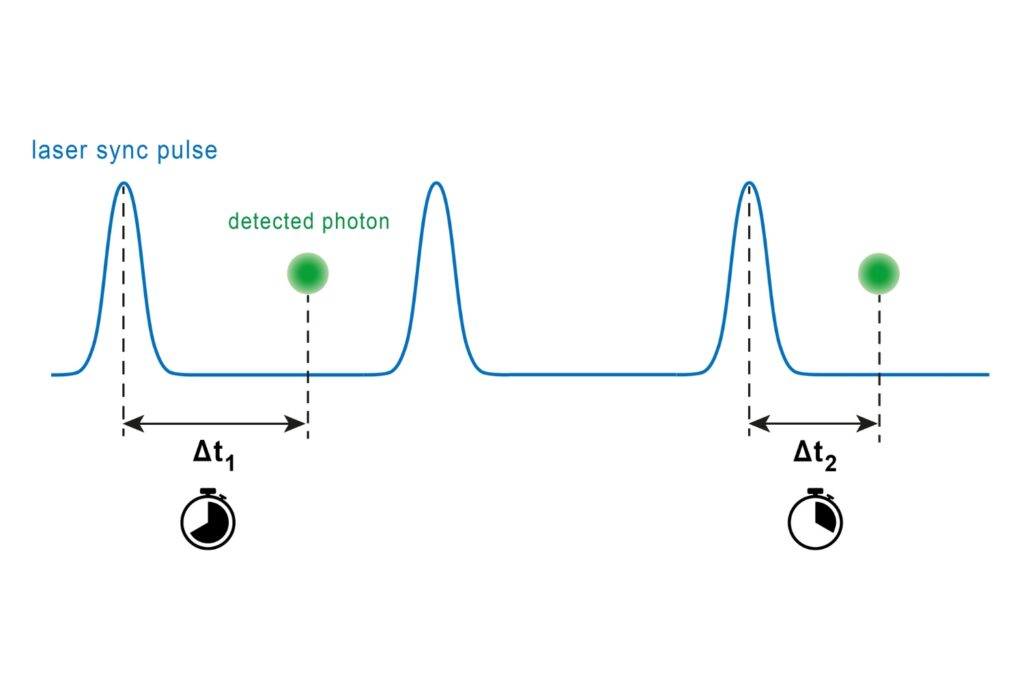
A photon timing technique that measures single-photon arrival times to resolve ultrafast dynamics in fluorescence, materials research, and quantum optics.
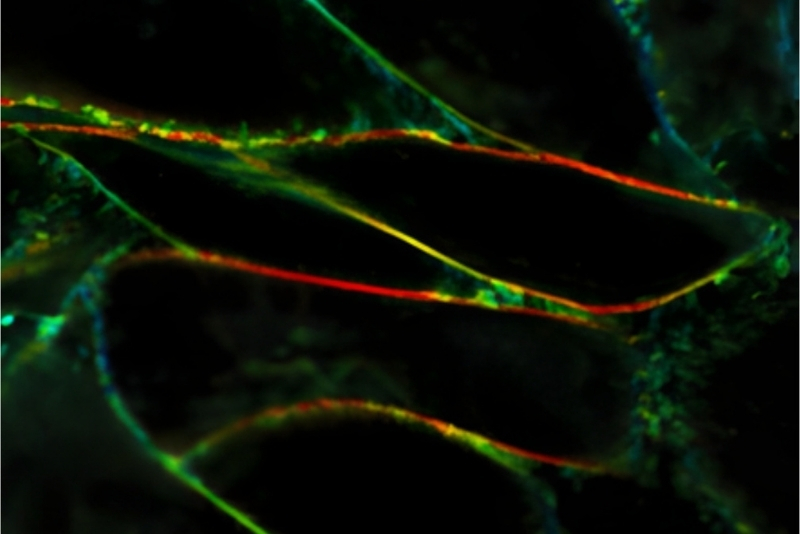
Fluorescence anisotropy can be implemented in both time-resolved spectroscopic measurements and polarization-resolved imaging to investigate molecular rotation, intermolecular interactions, and structural organization in biological systems.
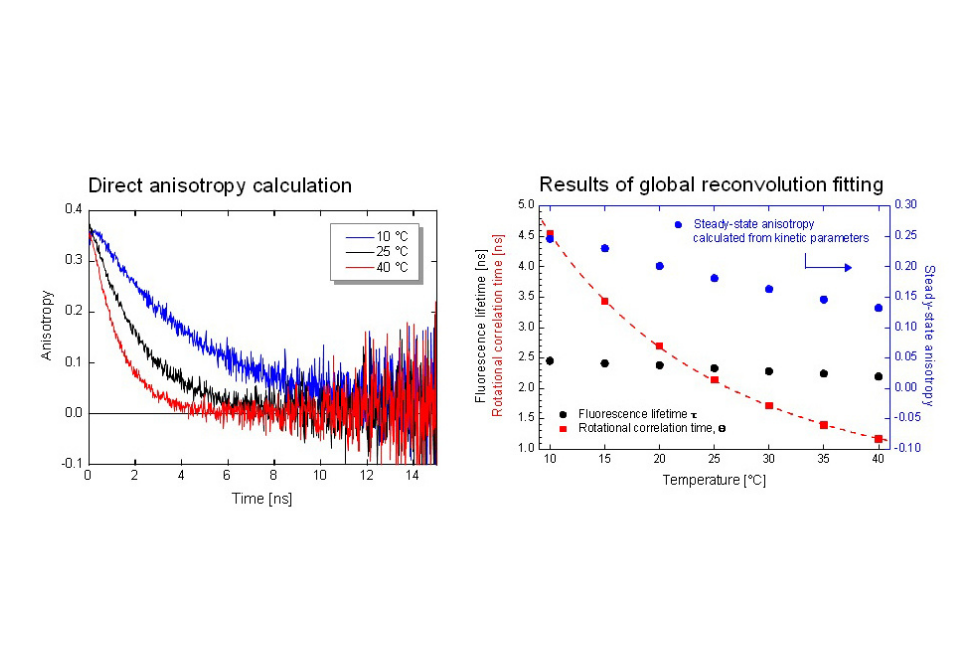
Time-resolved fluorescence anisotropy of Coumarin 6 was measured at different temperatures using the FluoTime 300 Photoluminescence Spectrometer. Global reconvolution analysis yielded rotational correlation times and fluorescence lifetimes. The results demonstrate temperature-dependent rotational dynamics consistent with viscosity changes and agreement with steady-state anisotropy predicted by the Perrin equation.
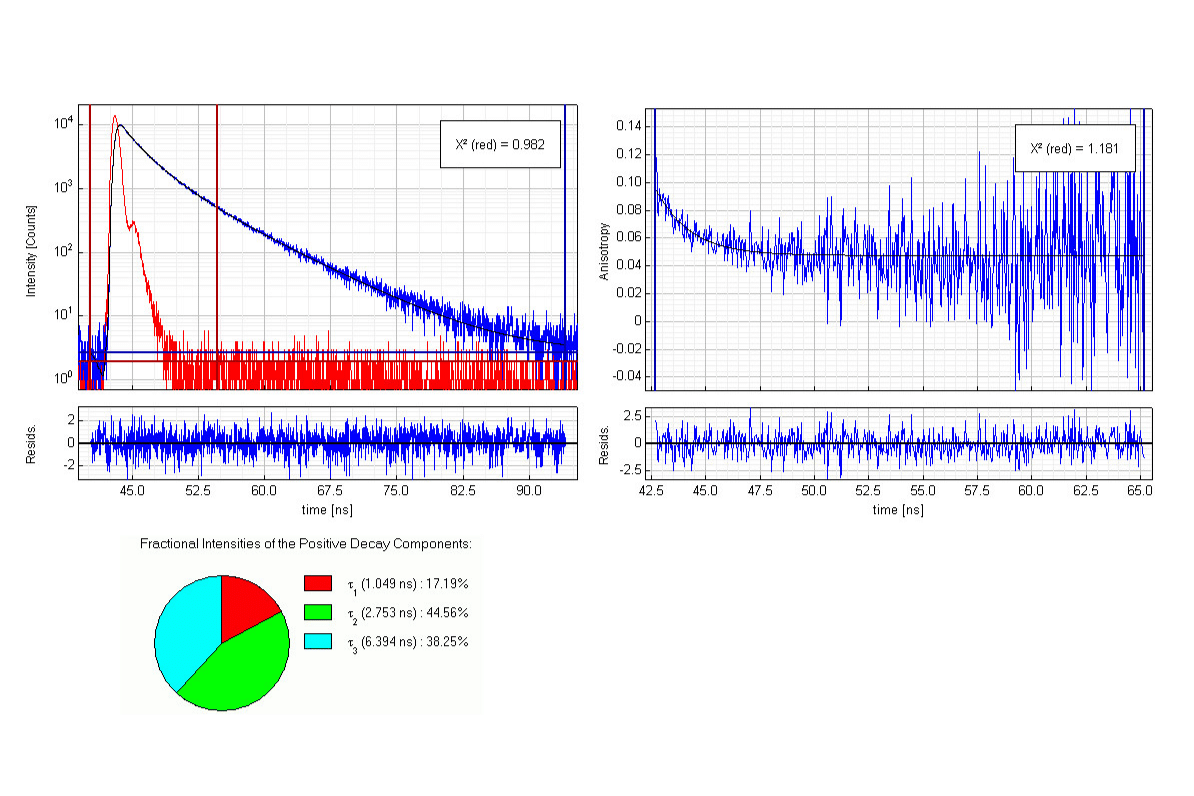
Time-resolved fluorescence anisotropy of apomyoglobin was measured following UV excitation at 280 nm. Polarized decay histograms enabled calculation of the anisotropy decay curve and determination of rotational correlation times. A residual anisotropy component indicated slower rotational processes beyond the nanosecond fluorescence lifetime window.
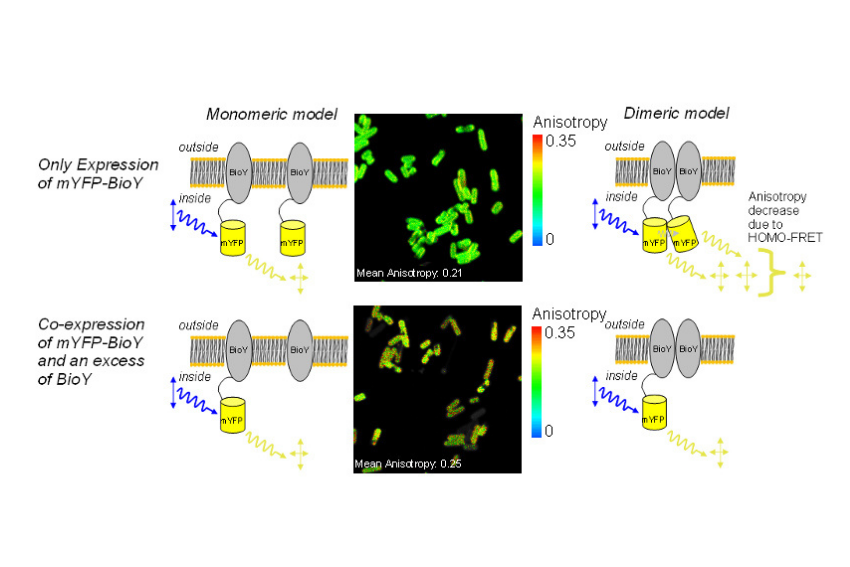
Fluorescence anisotropy imaging of mYFP-labeled BioY in E. coli was performed using a confocal microscope equipped with the LSM Upgrade Kit and polarization extension. Reduced mean anisotropy was consistent with HOMO-FRET, indicating dimer formation. Co-expression with excess unlabeled BioY increased anisotropy, supporting interaction between BioY subunits.
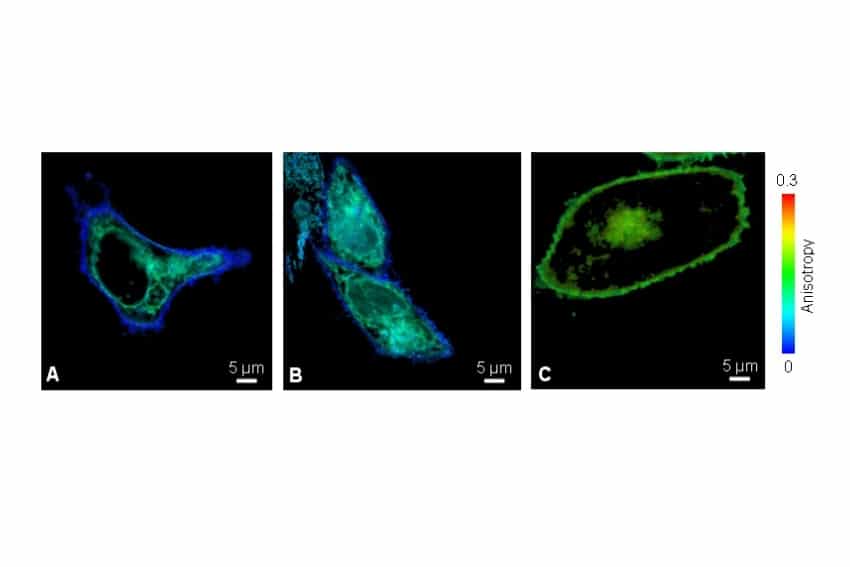
Fluorescence anisotropy imaging of v.m.p.-mYFP in living CHO cells was performed on a confocal microscope equipped with the LSM Upgrade Kit and polarization extension. Reduced anisotropy at the plasma membrane compared to intracellular regions was consistent with HOMO-FRET caused by clustering of the viral membrane protein.
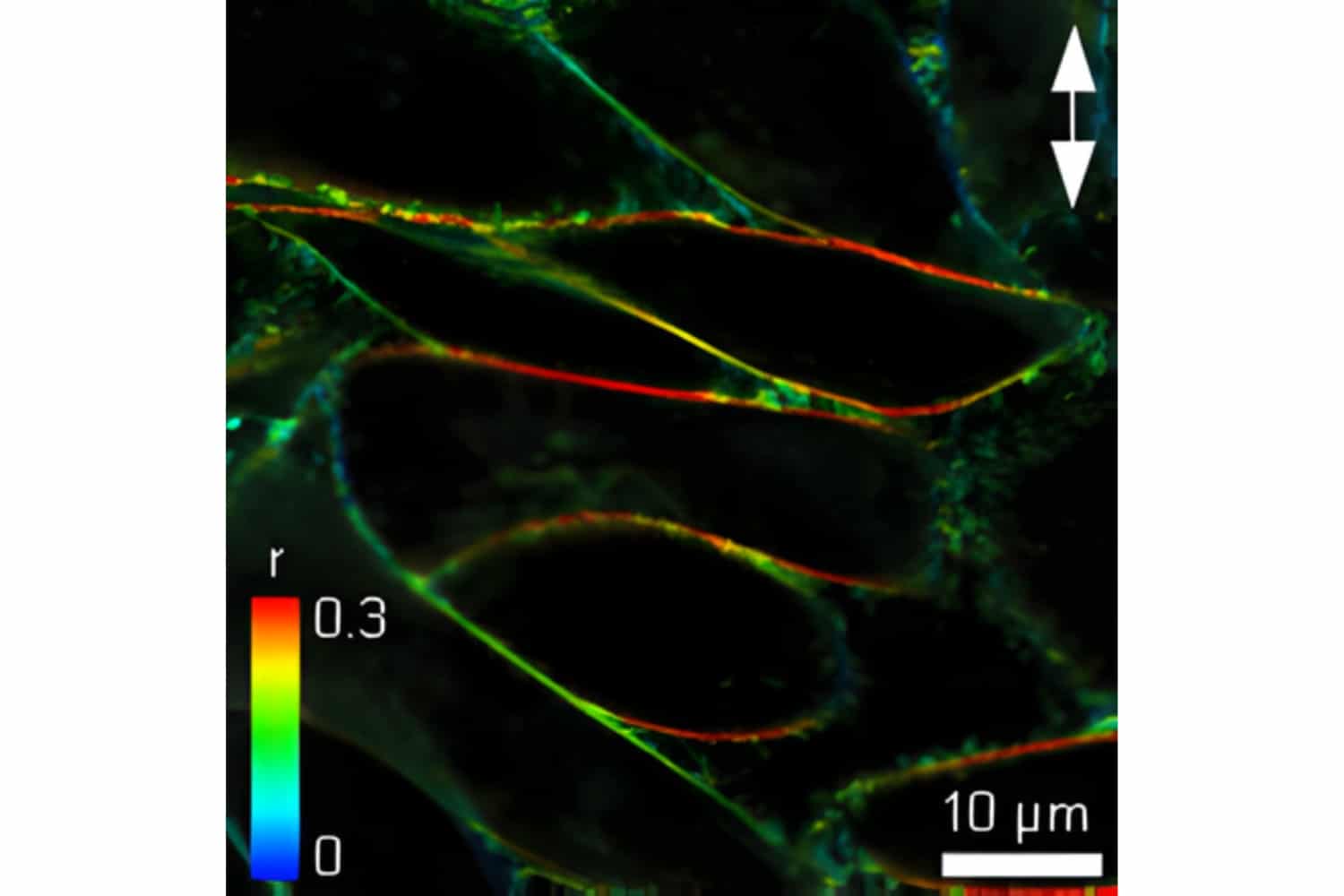
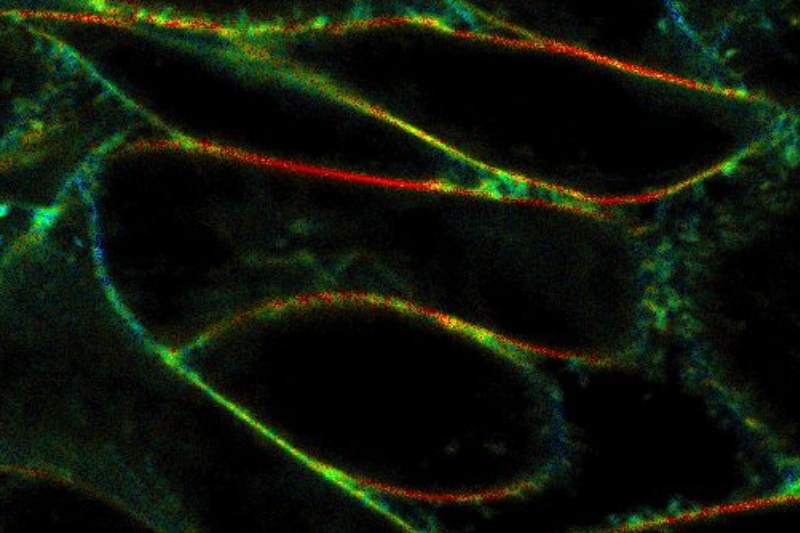
Fluorescence anisotropy imaging of CHO cells labeled with C6-NBD-PC was performed using an Olympus FluoView FV1000 equipped with the LSM Upgrade Kit and polarization extension. Higher anisotropy in horizontal membrane regions compared to vertical membranes is consistent with liquid-ordered membrane phases and restricted probe orientation.
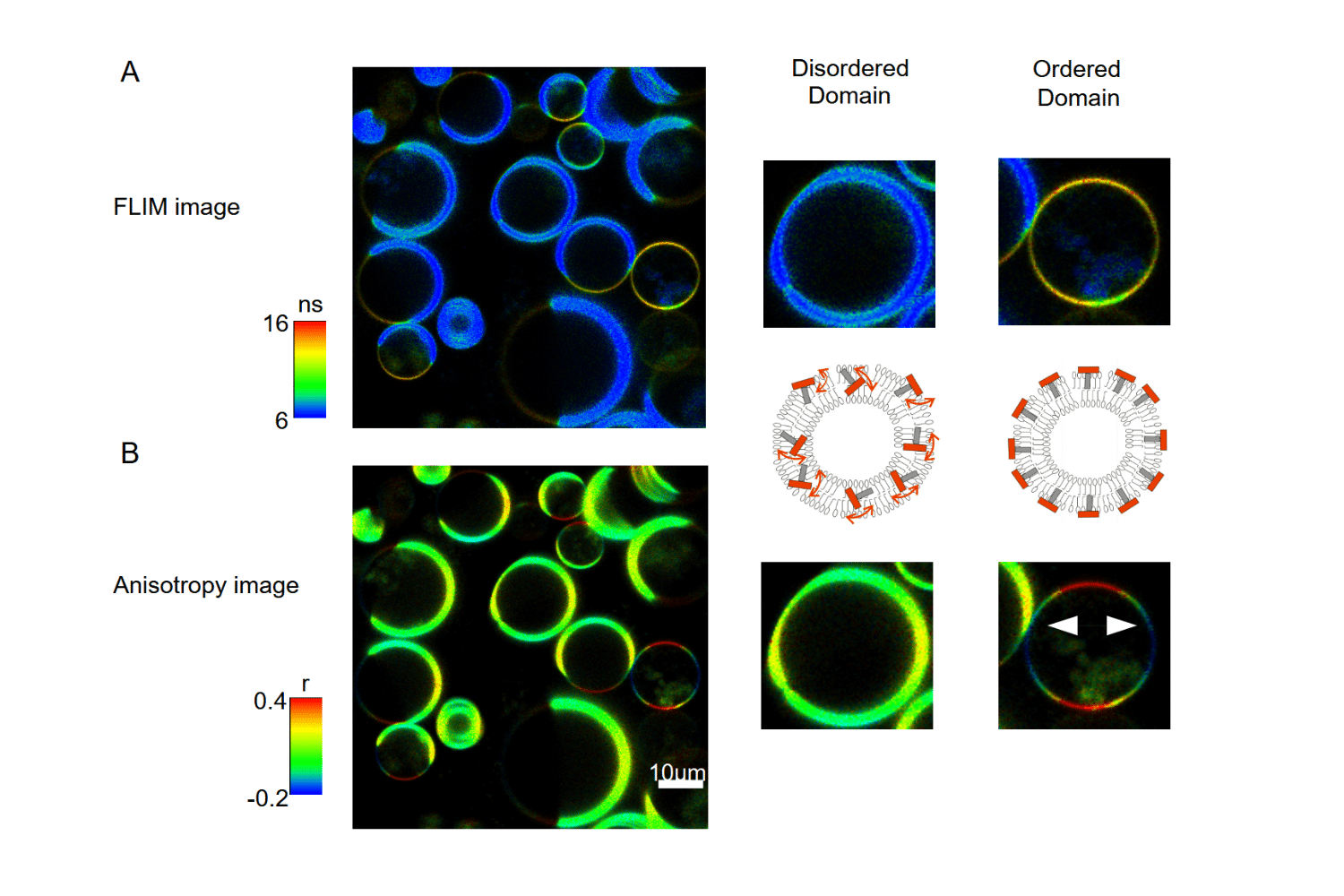
Fluorescence anisotropy imaging was used to distinguish liquid-ordered and liquid-disordered domains in giant unilamellar vesicles. Differences in polarization retention revealed variations in molecular orientation and rotational mobility across the membrane, enabling spatial mapping of structural heterogeneity at the micrometer scale.